商业广告QQ
896000434
896000434
m6A修饰是真核生物常见的mRNA修饰,已成为一种广泛的调控机制。m6A修饰由甲基化酶和去甲基化酶动态调控,甲基化酶主要包括METTL3、METTL14和WTAP等,去甲基化酶主要包括ALKBH5和FTO等。本研究中,作者们首先构建部分肺切除模型(Pneumonectomy,PNX),筛选出在肺再生内皮细胞中显著变化的METTL3,随后利用内皮条件性敲除Mettl3(Mettl3i∆EC/i∆EC)小鼠,发现内皮细胞Mettl3缺失导致肺再生中内皮细胞异常增殖,胶原纤维异常沉积,套叠式血管异常增多。为进一步在内皮细胞中研究METTL3调控的机制,作者们采用MeRIP测序联合纳米孔三代测序技术,证明了内皮细胞METTL3通过调控FOXO1的m6A修饰,在肺再生中维持内皮细胞的促再生Angiocrine功能。
作者们进一步发现,Mettl3i∆EC/i∆EC和Foxo1∆EC/i∆EC小鼠肺再生中异常增殖的内皮细胞为Rhoj+的细胞,且有Pillar Cell特征。这群异常增殖的内皮细胞通过增强血管微环境中促纤维化的Angiocrine信号,促进血管周围成纤维细胞的活化,导致肺再生向肺纤维化转变。在肺纤维化患者和肺纤维化小鼠模型中,作者发现肺内皮细胞RHOJ的表达显著上升。而在小鼠肺内皮细胞敲低RHOJ能够抑制异常的血管生成,减少促纤维化的Angiocrine信号,减轻肺纤维化。
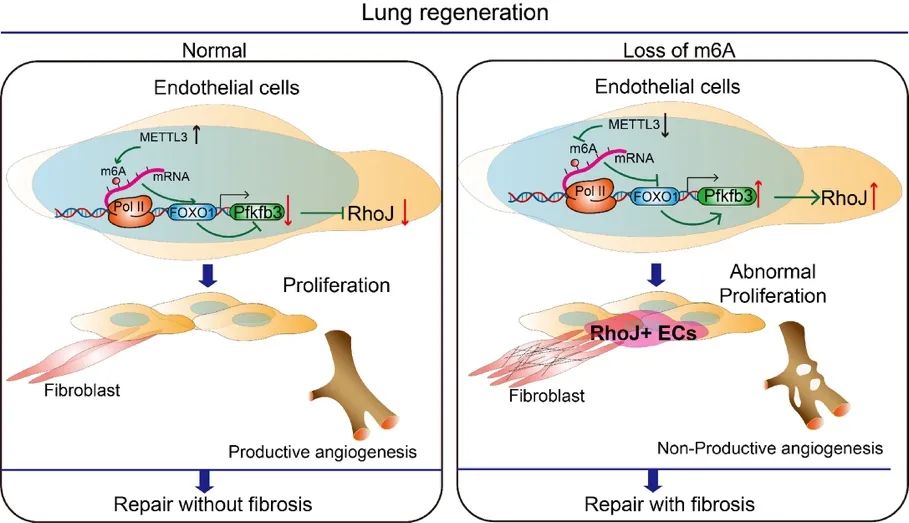
模式图(Credit:Science Translational Medicine)
总之,该研究发现了肺血管微环境中RHOJ+内皮细胞的形成机制,并揭示了在肺纤维化过程中RHOJ+内皮细胞组成的异常Pillar Cell可通过Angiocrine转变促进肺纤维化。所揭示的机制可为器官纤维化的再生疗法提供靶点,开发基于Angiocrine的药物。
